
頂端分裂組織の構造と機能の維持
2.1 植物の発生・成長と頂端分裂組織
双子葉植物の胚発生過程は、胚軸・幼根からなる体軸、体軸の両端に位置する茎頂・根端の頂端分裂組織、2枚の子葉、という基本的な体制(図10)を形成するのみである。われわれが目にする植物個体の地上部と地下部のほぼ全ての構造は、発芽後の "後胚発生" と呼ばれる過程で、頂端分裂組織(apical meristem)によって形成される。したがって、高等植物の発生・形態形成を理解する上で、頂端分裂組織についての理解は不可欠である。
これまで、茎頂分裂組織あるいは根端分裂組織に特異的な異常を示す突然変異体に着目して、(1) 胚発生過程における形成の機構、(2) 形成された後の構造と機能の維持機構、茎頂分裂組織、根端分裂組織のそれぞれについて独立に研究されてきた。しかし、茎頂分裂組織と根端分裂組織は外見上の違いはあるものの、共通の体制を持つと考えることができる(図10)。両方の頂端分裂組織で、共通の遺伝子が機能して、形成過程や構造・機能の維持に関与している可能性が考えられるが、そのような視点に基づいた研究があまりおこなわれていないのが実状であった。
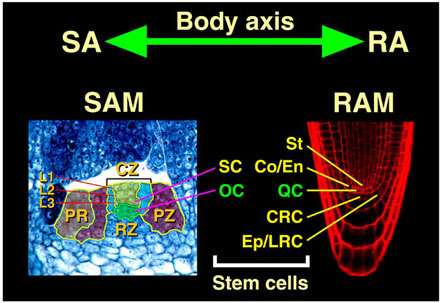
図10 高等植物の体制と頂端分裂組織
高等植物は、胚発生の過程で、体軸(body axis)の両端(茎頂と根端。図の SA, RA)に、茎頂分裂組織(shoot apical meristem, SAM)、根端分裂組織(root apical meristem, RAM)を形成する。
茎頂分裂組織には、組織学的・機能的に異なる領域がみとめられる。中央部には、central zone (CZ) と呼ばれる分裂活性が低い領域があり、その周囲を peripheral zone (PZ) と呼ばれる、活発に分裂し、器官原基(PR) を分化する領域が囲んでいる。CZ の下部には rib zone (RZ) と呼ばれる領域がある。こうした領域区分とは別に、外側から L1 (layer 1), L2, L3 という3種類の細胞系譜的に互いに独立の細胞層がみとめられる。L1, L2 はもっぱら垂層分裂(層面に対し垂直方向の分裂)をし、シロイヌナズナではそれぞれ1層からなる。これに対し、L3 の細胞分裂面はランダムで、第3層から内側の全ての細胞を含む。
根端分裂組織は、シロイヌナズナでは、非常に規則的な構造を持ち、4個の中心細胞(正中面では2個のみ見える)からなる静止中心(quiescent center, QC)の周囲に、5種類の始原細胞(Co/En, Ep, LRC, CRC, St)が位置する。始原細胞の数は一定している。
茎頂分裂組織では、CZ 中央部の第4層以下の少数の細胞(OCと表示)はWUSCHEL遺伝子を発現し、上層の細胞を幹細胞(stem cell)として維持する "organizing center" として機能する。一方、根端分裂組織では、QC がその周りの5種類の始原細胞(分化能が限定された幹細胞と考えることができる)の分裂と分化を制御し、"organizing center" として機能する。
2.2 FASCIATA遺伝子
われわれのグループは、そうした研究の現状を踏まえ、茎頂・根端の両頂端分裂組織の構造と機能の維持に共通して必要とされる遺伝子群を明らかにしたいと考え、まず、fasciata (fas)と呼ばれる、帯化突然変異(図11)に着目して研究を進めている。
fas突然変異体は、花茎が帯化する(茎の断面が円ではなく、扁平な帯状に広がる)突然変異体として、Reinholz (1966) により記載されたものであるが、後に、Leyser & Furner (1992) によって、帯化や葉序の不規則化に加えて、主根の長さも短くなることことが報告された。われわれは新規の遺伝子座に由来するものも含む多数のfas突然変異体を得て、その中から、FAS1, FAS2という2つの既知の遺伝子座に由来する変異を選び、まずこれらについて研究を進めた。

図11 茎の帯化を示すfas1変異体と野生型
野生型(右)とFAS1遺伝子を欠失した植物(左)。野生型に比べて、FAS1遺伝子を欠失した植物では茎の幅が広くなっている。中央の図はFAS1遺伝子を欠失した植物の花茎の先端部の拡大図。
発芽後の幼植物の頂端分裂組織の形態表現型の解析から、これまでに予想されていた茎頂分裂組織の構造異常(図12)に加え、根端分裂組織の構造異常も見出された(図13)。一方、成熟胚の頂端分裂組織の形態には明らかな異常はみとめられなかった(図14)。これらのことから、FAS1, FAS2遺伝子は形成後の頂端分裂組織の構造の維持に関わっている可能性が示唆された。
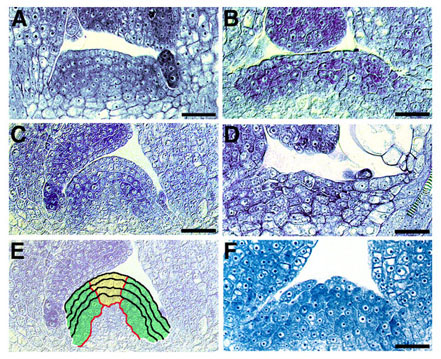
図12 fas1, fas2変異体と野生型の茎頂分裂組織
野生型(A, C。 Eに模式図を示す)、fas1変異体(B)、fas2変異体(D)、fas1; fas2二重変異体(F)。野生型でみられる、L1, L2 と L3の外層の整然とした細胞層の配列、CZ(Eの黄色の領域)と PZ(Eの緑色の領域)の明瞭な区別が、fas変異体では失われている。
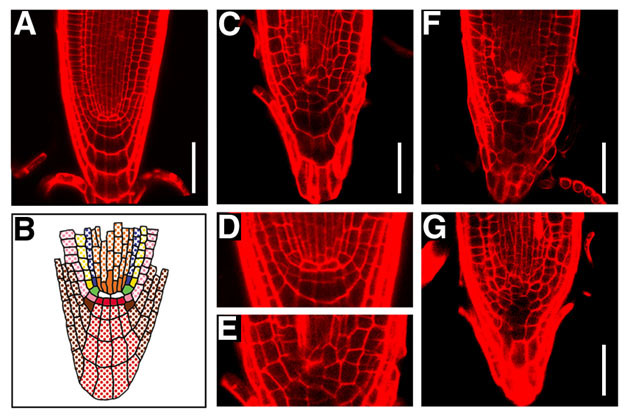
図13 fas1, fas2変異体と野生型の根端分裂組
See Cell 104(1), 131-142 (2001).
野生型(A, D)、fas1変異体(C, E)、fas2変異体(F)、fas1; fas2二重変異体(G)。野生型でみられる、規則的な細胞配列(Bに模式的に示す)が、fas変異体では失われている。静止中心(Bの白色で示した細胞)とそれを囲む各種始原細胞(Bで様々な色で塗りつぶして示した細胞)の配列の乱れは特に顕著である(D と E の拡大図を比較)。
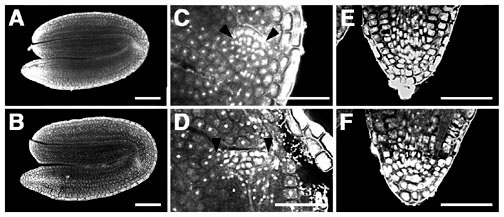
図14 fas1変異体と野生型の成熟胚
See Cell 104(1), 131-142 (2001).
野生型(A, C, E)、fas1変異体(B, D, F)。胚の全形(A, B)、茎頂分裂組織(C, Dで2つの矢尻の間の領域)と根端分裂組織(E, F)の拡大図。
T-DNA 挿入に伴う染色体欠失系統(Kaya et al. 2000)を用いて、FAS1遺伝子を同定したところ(FAS1遺伝子は、前述のFT遺伝子のすぐ隣の遺伝子であった)、ヒト、出芽酵母でこれまでに知られていたChromatin Assembly Factor-1 (CAF-1)の3つのサブユニットのうちの最も大きなサブユニット(ヒト、出芽酵母のp150、Cac1pに対応)と相同性を持つ蛋白質をコードすることが判った(Kaya et al. 2001)(図15)。CAF-1は複製後、あるいは除去修復後の DNA 上にヌクレオソームを形成するヒストン・シャペロンである。
われわれは、ヒト、出芽酵母のCAF-1が3つのサブユニットからなること、fas1, fas2突然変異体の表現型がよく似ており、また、二重変異体の表現型が単一変異体の表現型と区別しがたいこと、などから、FAS2遺伝子がCAF-1の残る2つのサブユニットのうちのひとつにあたるものをコードする遺伝子であると予想した。この予想は正しく、FAS2遺伝子はCAF-1の2番目のサブユニット(ヒト、出芽酵母のp60、Cac2pに対応)にあたるものであることが明らかになった(Kaya et al. 2001)(図15)。残念ながら、これまでに得ているfas突然変異体の中に、3番目のサブユニットに対応する遺伝子に欠損を持つものは見つかっていない(その後、別の研究グループによって、3番目のサブユニットの遺伝子AtMSI1の機能欠損変異体は致死になることが報告された)。
われわれは、米国コールド・スプリング・ハーバー研究所の B. Stillman 博士、柴原慶一博士(現・国立遺伝学研究所育種遺伝研究部門)との共同研究により、FAS1, FAS2 蛋白質がAtMSI1という3番目のサブユニットの候補蛋白質と複合体を形成し、複製に依存したヌクレオソーム形成というCAF-1としての機能を持つことを示した(Kaya et al. 2001)。 fas1, fas2突然変異体は、高等真核生物では今のところ唯一のCAF-1機能欠損変異体であり、CAF-1の生体内機能を明らかにする上で重要な手がかりを与えてくれると期待される。
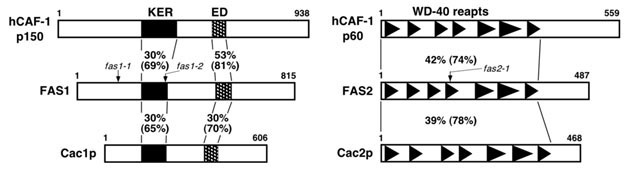
図15 FAS1, FAS2 蛋白質とそれぞれに相同を持つヒト、出芽酵母の蛋白質の比較
See Cell 104(1), 131-142 (2001).
FAS1 蛋白質(左の中段)と FAS2 蛋白質(右の中段)。FAS1 蛋白質は、ヒトの CAF-1 の p150 サブユニット(hCAF-1 p150)、出芽酵母の Cac1p と、FAS2 蛋白質は、ヒトの CAF-1 の p60 サブユニット(hCAF-1 p60)、出芽酵母の Cac2p と、それぞれ相同性を持つ。fas1-1, fas1-2, fas2-1の各突然変異(いずれもナンセンス変異)の位置を示す。
2.3 FASCIATA遺伝子を巡る今後の課題
fas1, fas2突然変異体は、CAF-1機能が欠損するものの致死とはならない(出芽酵母のcac1, cac2突然変異体も同様である)。しかし、シロイヌナズナのゲノム中には、FAS1, FAS2遺伝子と高い相同性を持つ遺伝子は存在しない。これらのことは、CAF-1の機能の一部を代行できるほかのヌクレオソーム構築因子が存在することを示唆する。われわれは、柴原慶一博士の研究室と協同して、そうした因子の解析を進めている。
fas1, fas2突然変異体では、CAF-1機能が欠損した結果、形成後の頂端分裂組織の構造が正しく維持されない。fas1, fas2突然変異体では、茎頂分裂組織におけるWUSCHEL (WUS) 遺伝子(図16)、根端分裂組織におけるSCARECROW (SCR) 遺伝子(図17)の発現制御にそれぞれ異常があり、これらが、頂端分裂組織の構造が安定に維持されない原因の一つであると考えられる(Kaya et al. 2001)。CAF-1が、DNA・クロマチン複製を通して遺伝子の発現状態を安定に維持することに関わっている可能性についても、検証していく予定である(成果の一端は、Ono et al. 2006 を参照)。
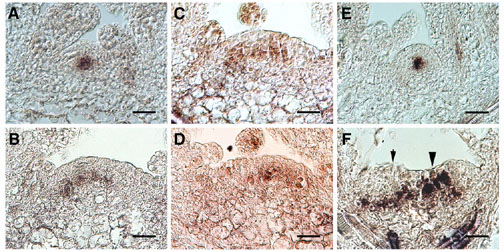
図16 fas1, fas2変異体と野生型の茎頂分裂組織におけるWUS遺伝子の発現
See Cell 104(1), 131-142 (2001).
野生型(A, E)、fas1変異体(B-D。同一の茎頂分裂組織由来の3つの切片)、fas2変異体(F)。濃褐色に染まっている細胞がWUS遺伝子を発現している。fas変異体では発現領域が顕著に広がっている。
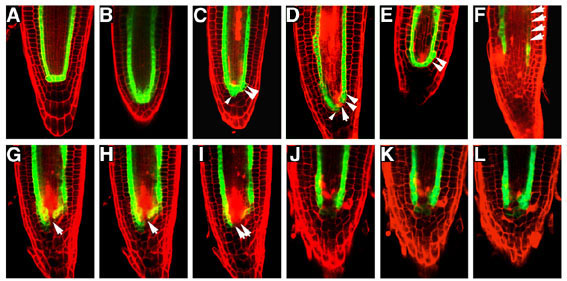
図17 fas1変異体と野生型の根端分裂組織におけるSCR遺伝子の発現
SCR遺伝子の発現は、SCR遺伝子プロモーターの制御下で発現させた緑色蛍光蛋白質(GFP)の蛍光(緑色)によって可視化している。野生型(A)、fas1変異体(B-L)。G-I と J-L は、それぞれ同一の根端由来の3つの光学切片像)。矢尻は異所的な発現を、矢印は発現の消失を、それぞれ表している。