
研究概要
植物が示す内的・外的環境へのさまざまな応答現象を研究する。2つのモデル植物を用いて、基盤をなす分子機構の解明と現象の解析を通して、陸上植物における基本原理の発見をめざす。主な研究項目として、以下の4つがある。
- 花成・生殖成長への移行を調節する分子機構に関する研究 (シロイヌナズナ・ゼニゴケ)
- フロリゲン (花成ホルモン) の輸送と作用の分子機構に関する研究 (シロイヌナズナ)
- 有性生殖 (特に配偶子形成) の制御機構に関する研究 (ゼニゴケ・シロイヌナズナ)
- 生活環の調節機構の進化に関する研究 (ゼニゴケ・シロイヌナズナほか)
研究内容
代表的な研究テーマ6つを紹介する。
- フロリゲンに関する研究 FT, FD (研究項目2)
研究室創設当初からの研究テーマで、これまでに成果の要点は図のようにまとめることができる。
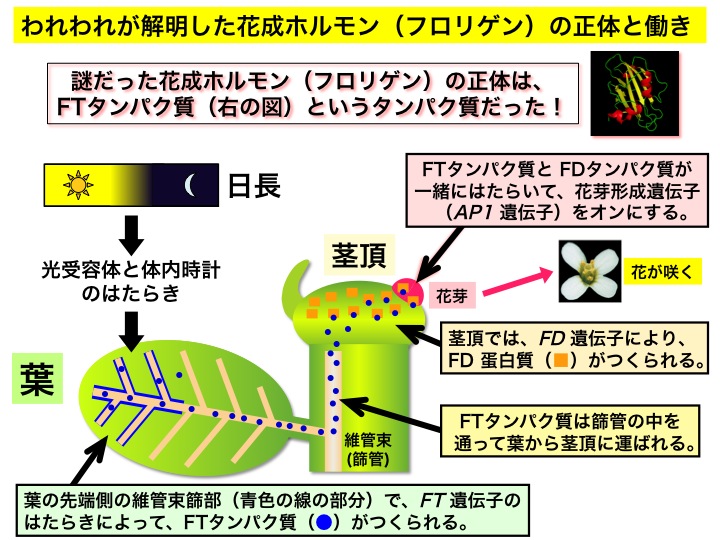
興味のある方は、京都大学の学外向け広報誌『紅萠』17号 (2010年3月発行) に掲載されている研究紹介をご覧になってください(p. 9-12)。 以下からダウロードできます。
『紅萠(くれなゐもゆる)』17号 PDF (3.45 MB)
- 日長応答機構の研究 phy, PIF, 概日時計 (研究項目1)
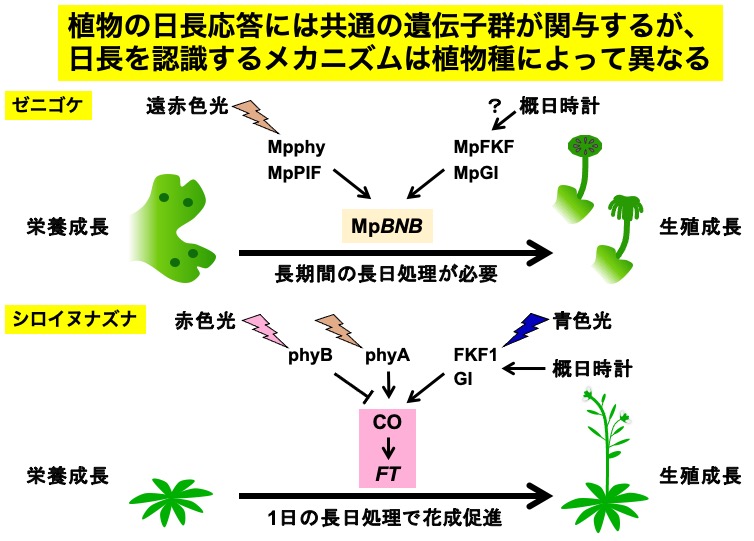
多くの植物は日の長さ(日長)を利用して季節を予測し、適切な季節に成長相転換を行なう。
シロイヌナズナやイネなどの被子植物は、光受容体と概日時計によって日長を認識している。
コケ植物ゼニゴケにおいても、共通の遺伝子群を用いて日長を認識することがわかった。一方
で、日長を認識するメカニズムはゼニゴケと被子植物で大きく異なることが予想された。
シロイヌナズナでは、 光受容体(phyA, phyB)や概日時計に制御される因子(GI, FKF1)
によって日長依存的にCO転写因子の発現や安定性が制御され、 CO転写因子がフロリゲン遺伝
子(FT )を制御することで成長相転換(花成)が誘導される。
これに対して、ゼニゴケでは、光受容体(Mpphy)は転写因子MpPIFとともに遠赤色光の
シグナルを伝達して生殖系列の決定因子である転写因子BONOBO (BNB) を制御する。また、
被子植物と共通してMpGI, MpFKFが日長依存的なBNB 遺伝子の制御に関わるが、このBNB 遺
伝子の制御に概日時計が関わるかは明らかではない。これらの遺伝子がどのように相互作用し、
日長依存的に BNB 遺伝子を制御するかを知ることが研究トピックスの1つである。
また、1回の長日処理で花成が促進されるシロイヌナズナとは異なり、ゼニゴケは10日以上
の長日処理が成長相転換に必要である。このことから、ゼニゴケには適切な長日条件の回数を
記憶する計数機構が存在すると考えられ、興味深い研究トピックスの1つである。
- 成長相転換に関わる保存された制御モジュールの研究 miRNA, SPL (研究項目1,4)
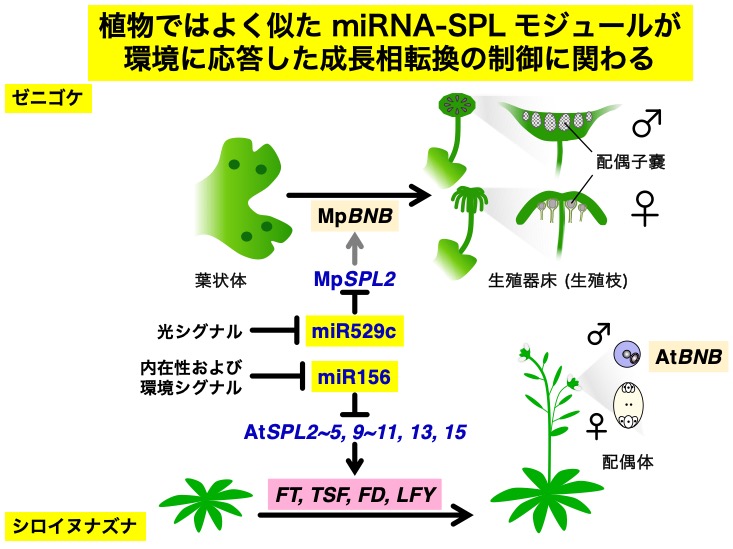
ゼニゴケ(コケ植物)とシロイヌナズナ(被子植物)は、どちらも長日条件のような環境条件に応答して成長相転換をおこない、生殖を開始する。
その過程の制御には、よく似た塩基配列を持つ miRNA とその制御標的となる SPL 転写因子が関わることがわかった。
シロイヌナズナでは、 SPL 転写因子はフロリゲン遺伝子(FT, TSF)やフロリゲン複合体の転写因子の遺伝子(FD)、花芽分裂組織遺伝子 LFY などを制御する。
これに対して、ゼニゴケでは、SPL 転写因子(MpSPL2)の下流には生殖系列の決定因子である転写因子BONOBO (BNB) の遺伝子があると考えられる。
MpSPL2 が BNB 遺伝子の発現制御にどのように関わるかは、現在の重要な研究課題の一つである。
また、光シグナル(前の項目)がどのようにして miR529c-SPL2 モジュールを制御しているのかを知ることはもうひとつの重要な研究課題である。
- 生殖系列の決定機構の研究 BNB (研究項目3,4)
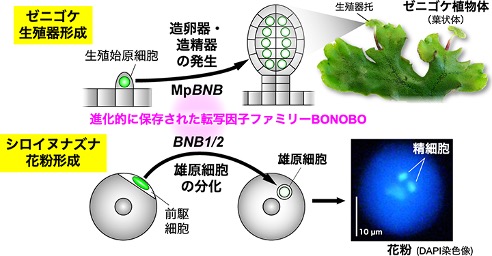
陸上植物の生殖細胞(卵・精細胞 [精子])は単数体(n)の多細胞組織である配偶体から分化する。 私たちがおこなってきたゼニゴケ変異体の分子遺伝学解析によって、配偶体である葉状体からの造卵器・造精器の始原細胞の分化と生殖器托の形成は、 マスター転写因子BONOBO (BNB)によってコントロールされていることが明らかになった。 分子系統樹によるとBNBは陸上植物全体で保存された転写因子であった。 さらにシロイヌナズナの解析から、被子植物のBNBオルソログは、雄性配偶体である花粉において雄原細胞の分化を制御することも明らかになった。 陸上植物において、少なくとも雄の生殖系列細胞の分化はBNBファミリーによって制御されていると考えられる。
詳しくはこちらを参照してください。
- 精細胞分化の制御機構の研究 MS1, DUO1
(研究項目3,4)
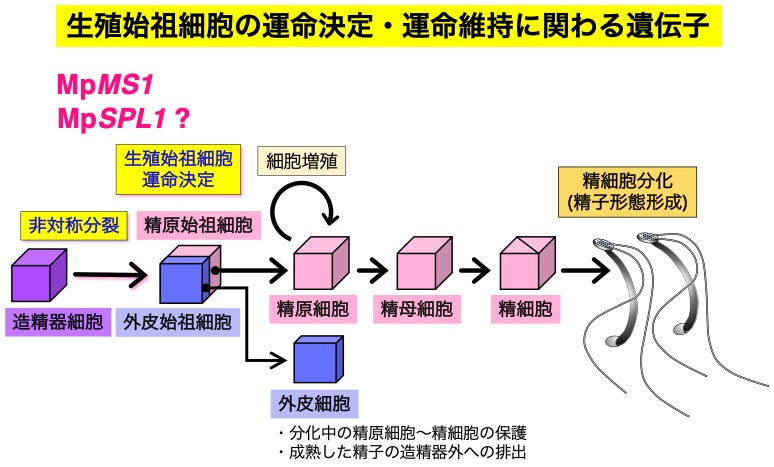
生殖器の発生過程の初期に、非対称分裂によって生殖始祖細胞と外皮始祖細胞の分化がおこる(図に示すのは雄の生殖器である造精器の例)。生殖始祖細胞は配偶子を分化し、
外皮始祖細胞は分化しつつある生殖細胞を保護する細胞層を形成する。造精器の場合には、外皮細胞は成熟した精子を排出する過程にも関わる。
現在進めている研究から、MpMS1 遺伝子が生殖始祖細胞と外皮始祖細胞の運命決定ないしは運命維持に重要な役割を果たすことがわかってきた。また、同様の役割は MpSPL1
にもあることが考えられる。
MpMS1 遺伝子はこの図に示す造精器だけではなく雌の生殖器である造卵器においても、
生殖始祖細胞と外皮始祖細胞の運命決定ないしは運命維持に重要な役割を果たしている。
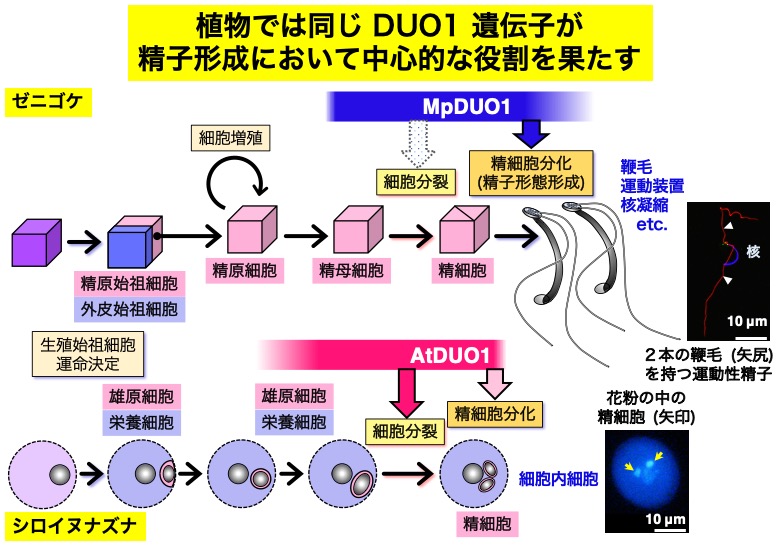
コケ植物(ゼニゴケ)と被子植物(シロイヌナズナ)の精細胞は、前者では2本の鞭毛を持つ運動性の精子であるのに対して、後者では花粉の中にある非運動性の「細胞内細胞」である。
分化の過程も、前者では一回の不等分裂、十数回の等分裂による増殖、そして最後の等分裂の後の精子形態形成と複雑であるのに対して、後者では一回の不等分裂、雄原細胞の取り込み、一回の等分裂と精細胞分化と簡略化されている。
そのような大きな違いにもかかわらず、同じ DUO POLLEN 1 (DUO1 ) という遺伝子が精細胞分化のマスター制御因子としてはたらく。さらに、2つの植物間で遺伝子に互換性がある。
2つの植物では、DUO1 遺伝子の制御下で異なる制御モジュールがはたらくことで、全く異なった形態と性質の精細胞が分化すると考えられる。そのような「制御モジュール」の同定が当研究室の大きな研究課題の一つである。
詳しくはこちらを参照してください。